0. 목차
- 대뇌피질
- '감각'의 메커니즘
- '기억'의 메커지즘
- '운동'의 메커니즘
- '사고'의 메커니즘
- 글리아 세포
1. 대뇌피질
'인간 게놈 프로젝트(HGP: Human Genome Project)'는 30억 개의 염기쌍으로 이루어진 DNA를 조사해서 어디에 어떤 유전자가 있는지를 밝히는 이른바 '지도 만들기'였다. 다음은 뇌다. 1000억 개가 넘는 뉴런들이 만들어내는 아주 복잡한 시스템을 알기 위해서는 우선 뇌의 지도를 만들어야 한다.
뇌과학에서 등장한 '비삽입 뇌영상법'이라 총칭되는 몇가지 방법은 사람들이 뇌에 접근하는 최대의 무기로 사용된다. 이러한 방법들을 사용해서 우리는 사람에 뇌에서 작용하는 영역을 가시화할 수 있다. 그 중 대표적인 것이 바로 'fMRI'라는 것이다. fMRI 장치를 사용하면 '칼럼'이라는 대뇌피질 미세구조의 활동을 포착할 수 있다.
인간의 대뇌는 좌우 2개로 나누어져 있다. 대뇌 반구의 표면에 자리하는 부위를 대뇌 피질이라고 한다. 대뇌피질에서는 수만 개의 뉴런이 다발을 이루어 원기둥 모양을 구조를 이루고 있다. 칼럼은 원기둥이라는 뜻이다. 이 원기둥의 지름은 약 0.5~1mm이고 높이는 2~3mm이다. 우리 인간 사회에서 사람들이 팀을 이루어 일을 하는 경우가 많은 것처럼, 대뇌피질의 뉴런도 하나의 칼럼이 하나의 팀으로 작용하는 것 같다. 대뇌피질에는 이러한 칼럼이 약 10만 개 정도 있으며, 대뇌 후두엽과 측두엽에서는 시각 정보를 인식하기 위해 칼럼이 하나의 모듈(기능 단위)로 작용하고 있다. 칼럼 하나하나의 기능을 따져 나가면, 칼럼 사이의 상호 작용 등 더욱 통합적인 정보 처리의 메커니즘을 밝혀낼 지도 모른다.
1-1. 대뇌 기능 지도 (브로드만의 뇌 지도)
독일의 해부학자인 '코르비니안 브로드만(Korbinian Brodmann, 1868~1918)은 대뇌 피질의 다양한 영역에 존재하는 뉴런의 형태를 조사하여, 그 차이에 근거해 대뇌 피질 각 영역에 1에서 52까지의 번호를 매겼다. 이후 1925년 '콘스탄틴 폰 에코노모(Constantin von Economo)'와 '게오르크 코스키나스(Georg N. Koskinas)'가 더 상세한 피질 지도를 발표하였다. 이후 브로드만의 뇌 지도(브로드만 영역; brodmann area)은 거의 한 세기 동안 토의와 논쟁을 거쳐 개선되었고 가장 잘 알려진 세포 구축 구조가 되었다. '코르비니안 브로드만'은 뉴런의 구성에 기초하여 정의한 영역을 피질의 다양한 기능과도 연관지었다.
예를 들어 브로드만 영역 1, 2, 3은 일차 몸감각피질이고, 영역 4는 일차 운동피질, 영역 17은 일차 시각피질, 영역 41, 42는 일차 청각피질과 밀접한 연관이 있다. 브로드만이 연관지은 것은 실제로 신경생리학, fMRI 등의 방법으로 확인한 브로드만 영역과 일치하였다. 예컨대, 왼쪽 브로드만 영역 44와 45는 말과 언어를 담당하는 '브로카 영역(Broca's Area)'과 일치하였다. 원래는 하나의 브로드만 영역이었던 것이 23a, 23b처럼 더 세분화되기도 했다. 지금도 fMRI 등에 의해 대뇌피질의 기능 지도 작동이 추진되고 있다.
아래에 그려진 '브로드만 뇌 지도(브로트만 영역)'은 두 개의 그림으로 구성되어 있다. 양 옆에서 보이는 표면을 '가쪽 표면(lateral surface)'이라고 하고, 좌우 반구 사이의 정분앙 분할면을 '가운데 표면'이라고 한다. 아래 그림의 '가쪽 표면'은 왼쪽 반구의 '가쪽 표면'이다.

1-2. 영장류의 브로드만 영역
| 영역 | - | 브로드만 영역 |
| 체감각 영역 | 감각 관련 피질 | 1, 2, 3, 5 |
| 시각 영역 | 감각 관련 피질 | 17, 18, 19, 20, 21, 37 |
| 청각 영역 | 감각 관련 피질 | 22, 41, 42 |
| 후각 영역 | 감각 관련 피질 | 34, 35, 36 ,38 |
| 다중 공간 영역 | 감각 관련 피질 | 7 |
| 시각, 체성, 청각, 언어 영역 | 감각 관련 피질 | 39, 40 |
| 번연 연합 영역 | 감각 관련 피질 | 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33 |
| 다중 내후각 뇌피질 영역 | 감각 관련 피질 | 28 |
| 뇌섭엽 피질 영역 | 감각 관련 피질 | 13, 14, 15, 16, 50 |
| 일차 운동 영역 | 운동 관련 피질 | 4 |
| 전운동 및 보조운동 영역 | 운동 관련 피질 | 6 |
| 전두시계 영역 | 운동 관련 피질 | 8 |
| 전전두 연합 영역 | 운동 관련 피질 | 9, 10, 11, 12, 44, 45, 46, 47 |
- 브로드만 영역 1: 일차 몸감각피질(primary somatosensory cortex)
- 브로드만 영역 2: 일차 몸감각피질(primary somatosensory cortex)
- 브로드만 영역 3: 일차 몸감각피질(primary somatosensory cortex)
- 브로드만 영역 4: 일차 운동피질(primary motor cortex)
- 브로드만 영역 5: 몸감각 연합피질(somatosensory association cortex)
- 브로드만 영역 6: 전운동피질(premotor cortex), 보조운동피질(supplementary motor cortex), (이차운동 피질(secondary motor cortex)
- 브로드만 영역 7: 몸감각 연합피질(somatosensory association cortex)
- 브로드만 영역 8: 전구엽 안구운동야(frontal eye fields)를 포함하는 영역
- 브로드만 영역 9: 등쪽가쪽 전전두피질(dorsolateral prefrontal cortex)
- 브로드만 영역 10: 앞쪽 전전두피질(anterior preforntal cortex)
- 브로드만 영역 11: 안와전두 영역(orbitofrontal area)
- 브로드만 영역 12: 안와전두 영역(orbitofrontal area)
- 브로드만 영역 13: 섬 피질(insular cortex)
- 브로드만 영역 14: 섬 피질(insular cortex) / 브로드만 14는 사람이 아닌 유인원에게만 있다.
- 브로드만 영역 15: 앞쪽 측두엽(temporal lobe)
- 브로드만 영역 16: 섬 피질(insular cortex)
- 브로드만 영역 17: 일차 시각피질(primary visual cortex, V1)
- 브로드만 영역 18: 이차 시각피질(secondary visual cortex, V2)
- 브로드만 영역 19: 연합 시각피질(associative visual cortex, V3, V4, V5)
- 브로드만 영역 20: 아래쪽 측두이랑(inferior temporal gyrus)
- 브로드만 영역 21: 가운데 측두이랑(middle temporal gyrus)
- 브로드만 영역 22: 위쪽 측두이랑(superior temporal gyrus) / 꼬리쪽 부분은 대게 '베르니케 영역'이 있는 곳으로 간주됨.
- 브로드만 영역 23: 배쪽 뒤쪽 띠다발피질(posterior cingulate cortex)
- 브로드만 영역 24: 배쪽 앞쪽 띠다발피질(anterior cingulate cortex)
- 브로드만 영역 25: 무릎밑 영역(subgenual area, 배쪽안쪽 전전두피질)
- 브로드만 영역 26: 대뇌 피질의 뒤뇌들보팽대(retrosplenial region) 중 바깥뇌들보팽대 부분(ectosplenial portion)
- 브로드만 영역 27: 조롱박피질(piriform cortex)
- 브로드만 영역 28: 배쪽 내후각피질(entorhinal cortex)
- 브로드만 영역 29: 뒤뇌들보팽대(retrosplenial region), 띠다발피질(cingulate cortex)
- 브로드만 영역 30: 띠다발피질(cingulate cortex)의 일부
- 브로드만 영역 31: 등쪽 뒤쪽 띠다발피질(posterior cingulate cortex)
- 브로드만 영역 32: 등쪽 앞쪽 띠다발피질(anterior cingulate cortex)
- 브로드만 영역 33: 앞쪽 띠다발피질(anterior cingulate cortex)의 일부
- 브로드만 영역 34: 해마곁이랑(parahippocampal gyrus)에 있는 등쪽 내후각피질(entorhinal cortex)
- 브로드만 영역 35: 후각뇌고랑(rhinal sulcus)의 후각주위피질(perirhinal cortex)
- 브로드만 영역 36: 현재 후각뇌고랑(rhinal sulcus)의 후각주위피질(perirhinal cortex)의 일부인 바깥후각 영역(ectorhinal area)
- 브로드만 영역 37: 방추형이랑(fusiform gyrus)
- 브로드만 영역 38: 위쪽 및 가운데 측두이랑(temporal gyrus)의 가장 입쪽 부분인 측두엽극 영역(temporopolar area)
- 브로드만 영역 39: 각이랑(angular gyrus) / 베르니케 영역의 일부로 간주됨.
- 브로드만 영역 40: 모서리위이랑(supramarginal gyrus) / 베르니케 영역의 일부로 간주됨.
- 브로드만 영역 41: 청각피질(auditory cortex)
- 브로드만 영역 42: 청각피질(auditory cortex)
- 브로드만 영역 43: 일차 미각피질(primary gustatory cortex)
- 브로드만 영역 44: 아래 전두이랑(inferior frontal gyrus)의 일부인 덮개부(pars opercularis) / 브로카 영역의 일부.
- 브로드만 영역 45: 아래 전두이랑(inferior frontal gyrus)의 일부인 삼각부(pars triangularis) / 브로카 영역의 일부.
- 브로드만 영역 46: 등쪽가쪽 전전두피질(dorsolateral prefrontal cortex)
- 브로드만 영역 47: 아래 전두이랑(inferior frontal gyrus)의 일부인 안와부(pars orbitalis)
- 브로드만 영역 48: 측두옆 가운데 표면의 일부인 해마이행부뒤 영역(retrosubicular area)
- 브로드만 영역 49: 설치류의 해마이행부곁 영역(parasubicular area)
- 브로드만 영역 50:
- 브로드만 영역 51:
- 브로드만 영역 52: 측두엽과 섬엽(insula)이 만나는 섬엽곁 영역(parainsular area)
2. 감각의 메커니즘
2-1. 시각 정보의 인식
우리가 사과를 보았을 때 어떻게 우리가 본 것이 사과라는 것은 어떻게 알 수 있을까? 이는 뇌과학에서 매우 중요한 문제이다.
- 망막에서 포착된 시각정보가 1차 시각령에 이른다: 눈에 들어오는 빛은 먼저 망막에 있는 시세포를 자극해 흥분시킨다. 그 흥분은 대뇌 후두엽의 '1차 시각령'에 이른다. 이 때 시야의 좌우는 반전된다. 즉, 왼쪽 시야는 주로 우뇌로 들어가고, 오른쪽 시야는 주로 좌뇌로 들어간다. 나아가 상하 방향도 반전된다.
- '1차 시각령'에는 '기울기 칼럼'과 '색깔 칼럼'이 있다: 대뇌 후두엽의 1차 시각령(V1)에서는 선분의 '기울기에 반응하는 뉴런'과 '빛깔에 반응하는 뉴런'이 각각 모여 칼럼 구조를 형성하고 있다. (후두엽의 시각령은 1~5차까지 복잡한 구조로 되어 있음) 우리가 보는 세계의 모습은 뇌 속에서 '어느 각도로 기울어진 아주 짧은 직선'과 '색깔'이라는 단 두 가지 요소로 분해된다. 또 왼쪽 눈에서 입력을 전문으로 받는 칼럼(초록색)과 오른쪽 눈에서 입력을 받는 칼럼(붉은색)이 줄무늬처럼 늘어서 있다. 특정 색깔에 반응하는 칼럼인 '블로브(blob)'도 군데군데 끼여 있다.
- 선분이 곡선으로 통합된다.: 이후 정보는 안쪽 시각 경로를 따라 측두엽으로 향하는 과정에서 물체의 윤곽이나 모양으로 재현된다. 후두엽 V4령에서는 윤곽을 구성하는 선분끼리 유연하게 연결되어, 기울기를 가진 곡선들이 통합되는 처리가 이루어지고 있음이 알려졌다.
- 측두엽 TE령 - 도형에 대응하는 칼럼이 반응: 정보는 결국 측두엽에 있는 TE령이라는 부위에 이른다. 이 경로의 앞쪽에 있는 TE령라는 부위에는 다양한 도형에 대응하는 여러 개의 칼럼이 있음이 원숭이의 실험으로 확인되었다. 하나의 칼럼은 영어로 말하면 26종류의 알파벳에 의해 표현되지만, 눈으로 본 것은 200종류 정도의 '도형 알파벳'의 조합으로 인식되는 것 같다. 우리는 닮은 것들을 '일반화'할 수 있다. 예를 들어 침팬치와 고릴라를 봤을 때 직감적으로 '원숭이'라고 일반화 할 수 있다. 'TE령'의 칼럼으로 말하면, 침팬지와 고릴라를 보았을 때 공통적으로 반응하는 몇몇 칼럼이 있다고 생각된다. 학습에 의해 그런 세트가 측두엽에서 만들어지는 것이 물체의 개념을 형성하는 메커니즘일지도 모른다.
2-2. 체성 감각 (자극에 대한 피부, 근육, 내부기관 등의 감각)
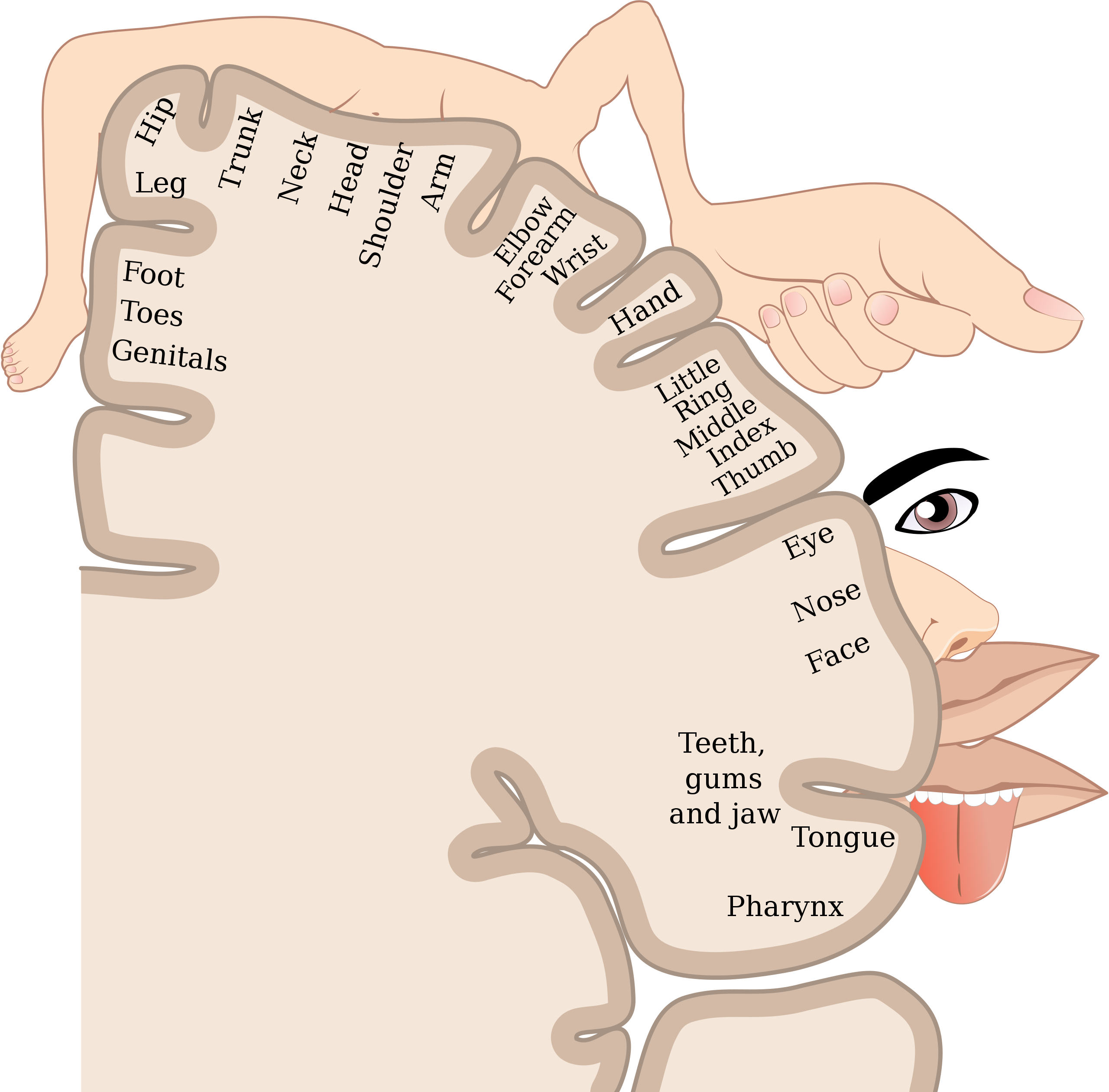
20세기 중엽 캐나다의 뇌외과 의사 '와일더 펜필드(Wilder Penfield)'는 뇌 수술을 받는 간질병 환자의 대뇌피질의 표면에 전기 자극을 주어 몸의 어느 부분이 만져지는 것처럼 느끼는지 조사했다. 그 결과, 몸의 각 부분에서 전해지는 촉각 자극이 '1차 체성 감각령'이라는 영역에 재현된다는 사실이 밝혀졌다. 1차 체성 감각령은 대뇌를 전두엽과 두정엽으로 나누는 큰 홈인 '중심구'의 바로 뒤에 있다. 그의 연구 결과를 그림으로 만든 것이 바로 '펜필드 호문쿨르스(Penfield Homunclus)'로, 이는 오늘날에도 유명하다. 참고로 '호문쿨루스(Homunclus)'는 '작은 사람'이라는 뜻이다. 아래의 그림은 '펜필드 호문쿨르스(Penfield Homunclus)'이다. 펜필드의 호문쿨루스에서는 얼굴과 손가락에 대응하는 1차 체성 감각령의 영역은 넓고, 각각의 부분이 촉각 자극 등에 민감하게 반응함을 나타낸다.
'펜필드 호문쿨루스'는 '1차 체성 감각령'이 체표면의 단순한 복사판인 듯하다는 인상을 오랫동안 많은 뇌 과학자들에게 남겨놓았다. '펜필드 호물쿨루스'를 보면 1차 체성 감각령에 있는 피질의 점 하나하나가 발이나 손, 얼굴 등의 대응하고 있는 것처럼 보인다. 하지만 펜필드가 실시한 방법은 지금 기준으로 보면 그다지 엄밀하다고 볼 수는 없다.
도호 대학의 의학부의 이와무라 교수는 촉각 자극과 1차 체성 감각령의 관계를 밝혀내기 위해 실험을 실시했다. 먼저 원숭이의 1차 체성 감각령에 전극을 넣고, 뉴런 하나하나가 어떤 몸짓 때 흥분하는지를 조사했다. ('집다', '쥐다', '더듬어 물체의 특징을 파악하다' 등...) 그 결과, 어떤 행동을 할 때 흥분하는 뉴런은 어느 위치에서 각각에 동작에 필요한 촉각 뉴런이 모여 기능적인 집단을 이루고 있었다. 또한 1차 체성 감각령의 뉴런 배치는 체표면의 단순한 복사판이 아님을 알 수 있었다. 뉴런 하나하나의 활동을 봄으로써 1차 체성 감각력과 체표면이 단순히 점 대 점 대응을 하는 것이 아님을 밝혀낸 것이다. 이런 연구에 의해 체표면의 단순한 복사판으로 간주되고, 정보 처리 능력이 없다고 생각되던 '체성 감각령'에 대한 고정관념이 바뀌었다.
3. '기억'의 메커니즘
3-1.기억과 '뇌 가소성'
- 스파인(Spine): 위에서 소개한 시각이나 체성 감각 등을 통해 얻은 정보는 뇌 속에 기억된다. 우리가 하나의 사항을 기억하면 수천 내지 수만 개에 이르는 시냅스에서 신호 전달의 용이함이 변한다고 한다. 시냅스에는 신호를 보내는 쪽의 뉴런인 '축삭 끝'과 신호를 받는 쪽 뉴런의 수상돌기에 있는 '스파인(spine)'라는 돌기가 미세한 틈을 두고 인접해 있다. 생쥐의 해마에 있는 뉴런을 조사하면, 스파인의 크기가 몇 분 단위로 활발히 변하고 있음을 관찰할 수 있다. 이 관찰을 통해, 강한 자극을 받은 시냅스일수록 스파인이 크게 부풀고, 그에 따라 신호도 전달되기 쉬워진다는 사실도 알아냈다. 이는 기억의 예비 과정이라고 할 수 있는 '시냅스 가소성(neuroplasticity)'이라는 현상을 시냅스 구조의 형태적 변화로 포착한 훌륭한 성과이다.
- 가소성: 그러면 여기서 말하는 '가소성'이란 무엇일까? 점토를 예로 들어 생각해 보자. 점토는 힘을 가하면 쉽게 변형하지만 그 변형된 상태를 그 뒤 계속 유지하는데, 이러한 성질을 '가소성'이라고 한다. 반면 금속은 상당을 힘을 가해야 변형시킬 수 있고, 고무 같은 소재는 힘을 가하고 있는 동안은 변형되지만 힘을 빼면 원상태로 돌아온다. 점토는 '가소성'을 가짐으로써 '힘이 가해졌다'는 경험을 기억할 수 있는 것이다. 우리가 겪는 경험 또한 시냅스가 가진 가소성에 의해 뇌안에 새겨진다. '기억(Memory)'은 크고 작은 다양한 스파인을 지닌 시냅스의 조합에 의해 뇌 안에 고정되어 있다.
3-2. '해마'와 기억 회로
대뇌의 측두엽 안쪽에는 '해마'라는 기관이 있는데, 이 해마는 새로운 기억을 획득하는 곳이다. 기억의 메커니즘을 분자 차원에서 알 수 있다면 기억력을 높이는 약이 현실화될 수 있다. 그리고 이러한 기억의 메커니즘을 파악하기 위해 실시한 실험 하나를 소개한다. 녹아웃 생쥐(유전자 재조합 기술에 의해 생성된, 특정 유전자가 결여된 생쥐)를 이용하면 해마의 기억 메커니즘에 대해 연구할 수 있다. 특정 유전자가 파괴된 생쥐의 학습 능력을 관찰함으로써, 그 유전자가 만드는 단백질 분자가 정말 기억에 필요한지를 알 수 있게 되는 것이다. 나아가 유전자가 파괴되는 영역을 한정시킴으로써, 해마 속의 어떤 뉴런 회로가 기억의 획득에 필요한지를 특정할 수 있다. 이 연구에서 기억의 획득에는 해마에 있는 CA1령이라는 영역의 '시냅스의 가소성'이 필요하다는 사실이 알려졌다. 'CA3령'이라는 영역이 '상기'에 중요하다는 사실도 알려졌다. 예컨대, 배우의 이름에서 영화의 한 장면을 떠올리는 것처럼 사소한 계기로 여러 가지 정경을 회상하는 경우에는 'CA3령'이 중요한 역할을 한다.
최근의 뇌 연구는 기억의 획득이나 상기뿐만 아니라 기억을 지우는 메커니즘에도 접근하고 있다. 대뇌 전두 연합령은 감정을 다스리는 대뇌 편도핵 등에 작용해, 좋지 않은 기억을 억제하고 있다. 그때 작용하는 분자 메커니즘은 알려졌다. 이미 미국의 제약회사에서 '끔찍한 과거의 기억을 억제하는 약' 개발에 들어간 것으로 보인다.
해마에서 만들어진 기억은 해마에 일시적으로 보존되지만, 최종적으로는 대뇌피질로 전송되어 측두엽의 시각령이나 청가력 등 그 정보를 최초로 처리하는 영역에 저장되리라고 생각된다. 이 전송은 주로 수면 중에 이루어진다는 학설이 있었다. 최근의 연구에서는 낮에 학습에 의해 만들어진 해마 뉴런의 흥분 패턴이 수면 상태의 생쥐 해마에서 '재연'되고 있는 모습이 관찰되었다. 이 재연은 수면 상태에서 이루어지는 기억의 전송이 포착된 것이다. 우리가 자면서 꾸는 꿈은 해마에서 만들어진 기억이 대뇌피질에 있는 기억의 창고에 정리되는 과정일지도 모른다.
4. '운동'의 메커니즘
4-1. 무의식 운동
신체에 명령을 내리기 위해 모든 행동에 의식적으로 판단할 필요는 없다. 우리가 달리기를 할 때의 뇌를 생각해 보자. 우리는 달리면서 '저 가로등 앞까지만 달리자', '피로하니 좀 천천히 달리자'같은 생각을 할 수 있다. 하지만 '우선 왼쪽 다리의 장딴지 근육에 힘을 주고 지면을 차면서 오른쪽 팔을 앞으로 흔들고...' 같은 생각은 할 필요가 없다. 일단 달리기 시작하면 자동적으로 근육이 균형 있게 움직이기 때문이다.
- 1차 운동령: 팔이나 다리에 있는 근육은 척수에서 뻗어 나온 운동 뉴런에서 온 자극에 의해 수축된다. 척수와 운동 뉴런 사이에 시냅스를 만들어 제어하고 있는 것이 대뇌 두정엽의 '1차 운동령'에서 축삭을 뻗은 뉴런이다. 1차 운동령의 작용은 의식되지는 않지만, 몸을 움직이는 동안 내내 왕성하게 이루어지고 있다. 1차 운동령의 뉴런은 칼럼 구조를 이루고 있다. 그래서 1차 운동령의 지령은 칼럼을 기능단위로 수행된다. 하지만 손가락의 운동을 생각할 경우 손가락 하나하나가 하나의 칼럼에 대응하는 것은 아니다. 하나의 칼럼은 여러 손가락에 지령을 내리고 있다. 예컨대 어느 칼럼은 엄지와 검지를 구부리는 운동을 지령하고, 어느 칼럼은 검지와 중지를 구부리는 운동을 지령하는 데 쓰인다.
- 고차 운동령: 우리가 하는 운동은 대뇌 전두엽의 '1차 운동령'에 있는 뉴런이 근육에 지령을 보냄으로써 이루어진다. 그 지령의 내용은 1차 운동령의 상위에 있는 '고차 운동령'이라고 총칭되는 전두엽의 영역이 기획, 구성한다. ('고차 운동령'에는 '운동 전령', '보조 운동령', 전보조 운동령', 대상피질 운동령' 등이 있다) 그를 위해 자신의 외부 세계에 대한 정보를 대뇌 연합령에서 받고 체내 상태에 대한 정보는 대뇌 번연계에서 얻는다. 이 정보를 바탕으로 상황에 따른 운동을 하도록 1차 운동령에 지령을 보내고 있는 것이다.
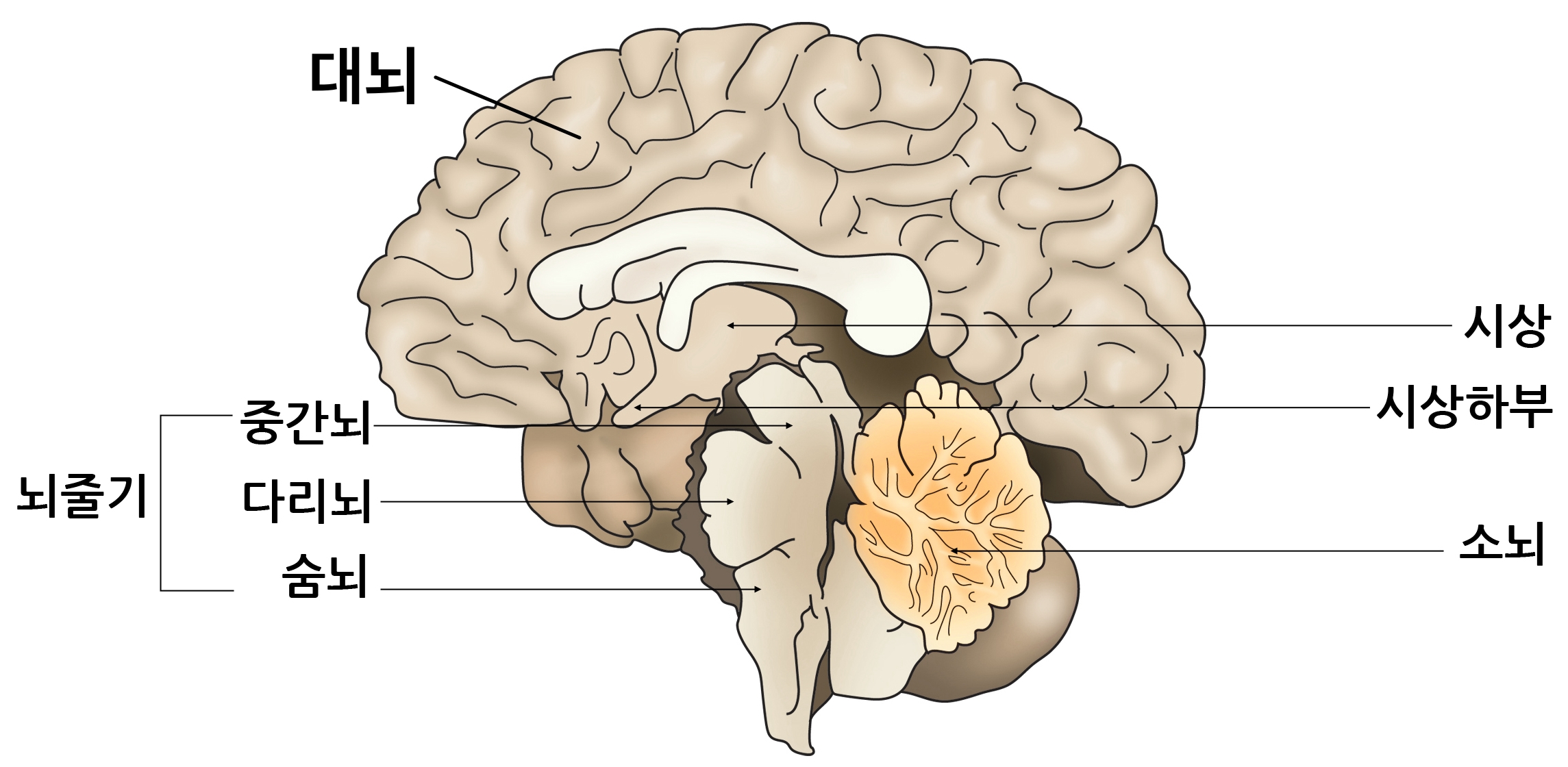
- 하부 조직: 대뇌 기저핵과 소뇌는 동작 조절을 위한 하부 조직으로 작용한다.
4-2. 소뇌와 '운동의 숙련'
처음 스키를 타는 사람은 아무리 스키에 대한 책을 읽어도 결코 제대로 탈 수 없다. 스키장에서 직접 타보면서 몸으로 익히는 수밖에 없다. 자전거를 타는 것도 그렇고, 수영도 마찬가지다. 왜냐하면 몸으로 익히는 이런 운동의 기억은 '대뇌'가 아니라 '소뇌'의 몫이기 때문이다.
- 소뇌는 '지우기'로 기억한다.: 소뇌 기억의 메커니즘은 흥미롭게도 대뇌의 해마에서 작용하는 메커니즘과 정반대이다. '해마'의 기억이 '쓰기'라면 '소뇌'의 기억은 '지우기'에 의해 이루어진다. 우리의 운동에 실수가 생기면 쓸데없는 운동을 유도하는 소뇌의 시냅스가 회로에서 사라져, 남은 시냅스만이 숙련된 움직임을 실현한다. 대뇌에서 온 신호는 평행 섬유를 타고 시냅스틀 통해 '푸르킨예 세포(Purkinje cell)'로 전달된다. 운동이 숙련되기 전에는 많은 시냅스가 효율적으로 전달한다. 하지만 스키를 타다 넘어지는 등의 실수를 하면 오류 신호가 상향 섬유를 통해 푸르킨예 세포로 전달된다. 그러면 그때 활동하고 있던 평행 섬유와 푸르킨예 세포 사이의 시냅스에서 전도 효율이 뚜렷하게 낮아진다. 이 현상을 '장기 억압'이라고 한다. 운동 실수의 원인이 된 시냅스는 이렇게 해서 회로에서 지워지고, 오류 신호를 만나지 않은 시냅스만 회로에 남아 숙련된 움직임을 실현한다.
- 소뇌의 '사고의 숙련': 소뇌은 운동뿐 아니라 사고의 숙련도 맡고 있다. 소뇌는 학습 기능에 의해 다른 회로의 작용을 복사할 수 있는 능력을 지니고 있다. 이 능력을 사용하여 소뇌는 대뇌의 사고 회로를 복사할 수 있다. 대뇌에서 이루어지는 사고는 의식으로 이루어진다. 우리는 보통 다른 연합령에서 만들어진 이미지를 대뇌 전두 연합령이 조종함으로써 사고한다. 하지만 똑같은 사고를 반복하면, 대뇌에 있는 이미지가 소뇌로 복사된다. 이렇게 복사된 것을 '내부 모델'이라고 한다. 예컨대 3X3이라는 계산 같은 경우에는 생각하지 않아도 답이 저절로 나온다. 이런 사고를 소뇌가 대신하고 있을지도 모른다는 것이다.
- 대뇌와 소뇌 사이의 연락 경로: 몸을 움직이는 지령은 대뇌 1차 운동령에서 뻗은 뉴런이 척수로 전해지지만 '교핵(橋核)'을 경유해 소뇌피질에도 전해져 처리된다. 소뇌에서의 출력은 척수로 향해 운동을 조정한다. 소뇌에서는 대뇌 쪽으로도 출력되고 있으며, 대뇌와 소뇌 사이에는 정보 경로가 만들어져 있다. 소뇌피질에서는 '올리브핵'이라는 곳에서도 입력이 이루어진다.
5. '사고'의 메커니즘
5-1. 전두 연합령
대뇌 전두엽 앞쪽에 자리하고 있는 '전두 연합령'은 '뇌속의 뇌'라고 말할 수 있는 최상위의 중추에 해당한다. 대뇌 전두 연합령은 영장류에서 특히 크게 발달한 부분이다. 붉은털 원숭이나 일본원숭이의 전두 연합령이 대뇌피질에서 차지하는 비율은 약 12%인데 비해, 사람의 경우는 약 30%나 된다. 전두 연합령은 영장류 이외의 포유류에서는 거의 발달되어 있지 않다. 고양이의 경우 전두 연합령이 대뇌피질에서 차지하는 비율은 2~3%밖에 되지 않고, 쥐는 전두 연합령이 아예 없다.
그런데 만약 전두 연합령이 파괴되면 어떻게 될까? 실제 그런 사례가 있다. 1848년 미국의 공사 현장 감독이던 '피니어스 게이지(Phineas Gage)'는 화약 폭발로 날아온 철봉이 머리에 꽂혀 전두 연합령의 대부분을 잃어버렸다. 그는 기적적으로 목숨은 건졌고 시각, 운동 능력 등에도 거의 장애가 없었지만 성격은 성격은 난폭해졌고 미래에 대한 계획성이나 꿈도 전혀 가질 수 없게 되었다. 각성제나 마약의 다수는 '전두 연합령'에 영향을 주어 인격을 붕괴시킨다. 전두 연합령은 바로 '사람을 사람답게 하는 뇌'라고 말할 수 있다.
전두 연합령에는 뇌의 여러 영역에서 아주 많은 입력과 출력이 이루어지고 막대한 양의 정보처리가 이루어진다. 그리고 그 정보 처리의 기능 단위는 칼럼 구조라고 생각된다. 칼럼 하나하나가 집적 회로 같은 역할을 하고, 그들이 모여 계층적인 네트워크를 이루고 있을 것으로 생각된다.
- 워킹 메모리(작업 기억): 전두 연합령에는 단기 기억의 일종인 '워킹 메모리(작업 기억)'라는 기능이 있다. 하지만 '워킹 메모리'는 단기 기억에는 없는 중요한 특징이 있는데, 바로 '행동을 위해서 쓰이는 기억'이라는 점이다. 예를 들어 가위를 찾기 위해 '가위'에 대한 기억을 워킹 메모리에 넣어 두었다가 옆방에서 그 기억을 사용해 찾는 것이다.
- 뇌 전체의 총괄과 감정 제어: 전두 연합령은 눈이나 귀 등을 통해 얻은 정보나 몸의 내부 환경(현재에 대한 정보), 미래의 예정(미래에 대한 정보), 기억이나 지식(과거에 대한 정보) 등에 접근할 수 있다. 그 막대한 정보 중에서 자신에게 의미 있는 정보만을 고르는데 이것을 '선택적 주의'라고 한다. 그리고 '워킹 메모리' 기능을 이용해 그것을 일시적으로 보존해 둔다. 그리고 그 정보를 바탕으로 행동이나 감정을 적절히 조절한다. 전두 연합령은 워킹 메모리를 중심으로 뇌 전체를 제어하는 통제 센터인 것이다.
5-2. 직감적인 결정
우리의 뇌는 직감적으로 일을 처리할 수 있다. 다만 직감적인 해결 방법은 철저하지는 못한 것이다. 직감적인 해결은 100점 만점의 답은 찾는 일은 거의 불가능하지만 대신 매우 짧은 시간에 90점의 답은 찾을 수 있다. 이런 직감적인 결정 능력은 왜 필요할까? 만약 적에게 습격을 당했을 때는 도망가는 경로를 생각하는 등의 치밀한 생각은 뒤로하고, 가능한 한 빨리 행동을 결정해야 한다.
- 디지털 컴퓨터: 컴퓨터는 디지털 정보만을 다루고 클록(clock; 일정 간격의 신호)'에 의해 '동기(신호 주파수 혹은 위상이 비교 신호의 그것과 어느 정상적인 관계에 있는 것을 말함)'되어 움직인다. 또 '처리 1을 수행한 다음에 처리 2를 한다.' 이러한 처리 방식을 '직렬 처리 방식'이라고 한다.
- 생물의 뇌: 우리의 뇌는 디지털과 아날로그를 여러 차원으로 나누어 사용하고 있다. 뉴런의 상태는 흥분의 유무만 본다면 디지털이지만, 그 흥분을 만들어내는 결정적인, 뉴런의 세포막이 나트륨 이온이나 칼륨 이온을 어느 정도로 통하게 하는가에 대한 정보는 아날로그다. 뉴런에 흥분이 생길 때의 간격도 아날로그다. 이처럼 디지털 정보와 아날로그 정보를 교묘히 뒤섞고 있는 상태를 카오스적이라고 말한다. 생물의 뇌는 카오적이다. 또 각각의 뉴런 활동에 동기성이 나타날 때도 있고, 그렇지 않을 때도 있다. 뉴런들의 정보 처리는 병렬 분산적으로 수행된다. 뉴런의 응답이 '카오스'적인 거동이라는 것은 실험으로 이미 증명되었다. '카오스(chaos)'란 결정론적인 법칙에 따르고 있음에도 불구하고, 그 결과가 불가능한 형태로 요동치는 성질을 말한다. 예컨대, 흔들이의 궤적은 단순하지만, 흔들이 2개를 이으면 그 움직임은 예측 불가능해져 카오스적이 된다. 이 카오스 상태는 아날로그 정보에서밖에는 생기지 않는다. 어쩌면 뇌가 내리는 직감적인 판단은 아날로그적인 성질이 초래하는 카오스에 본질이 있는 것일 지도 모른다.
- 카오스 컴퓨터: 아이하라 교수는 도쿄 전기대학 공학부의 호리오 요시히코 교수와 동동으로 카오스적인 거동을 기본 원리로 삼은 컴퓨터 칩도 개발하였다. 이 칩은 예를 들면 '방문 세일즈맨 문제'같은 조합 최적화 문제를 풀 때 위력을 발휘한다. 만약 이런 문제를 풀 때 일일이 모두 계산하여 문제를 풀려고 하면 엄청난 시간이 걸린다. 카오스 컴퓨터는 이러한 문제에 직감적으로 달려드는, 마치 뇌와 같은 컴퓨터라고 할 수 있다.
6. 글리아 세포
뇌의 기능은 뉴런과 시냅스에서 일어나는 일의 조합으로 설명할 수 있다. 그런데 이만큼 천문학적인 수의 요소로 이루어진 뇌라는 시스템에, 폭주 현상도 일어나지 않고 안정적일 수 있는 이유는 무엇일까? 뉴런으로 구성된 시스템을 감싸듯 하면서 조절하는 무언가의 또 다른 시스템이 있는 것은 아닐까?
전 세계의 뇌 연구자들이 관심을 받고 있는 뇌 안의 세포, '글리아 세포(Glia Cells)'라는 것이 있다. '글리아 세포'의 수는 뉴런의 10배 가까이 된다. 하지만 글리아세포는 뉴런처럼 전기적 흥분을 일으키지는 않는다. 그래서 글리아세포는 뉴런을 작용을 돕지만, 뇌 속에서 정보를 전달한 능력은 없는 것으로 여겨져, 오랫동안 크게 관심을 받지 못했었다. 하지만 글리아세포는 지금 크게 주목받고 있다.
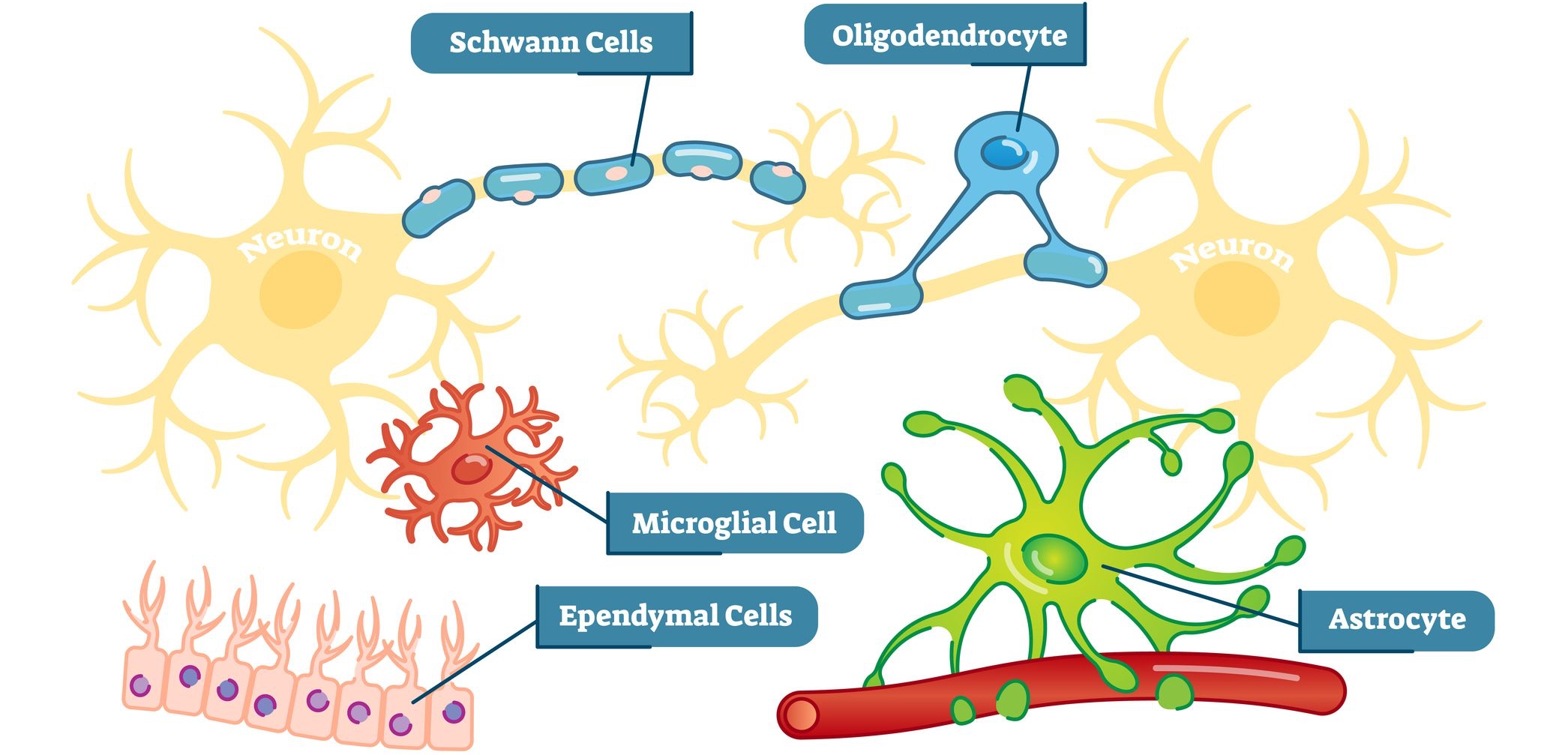
6-1. 성상교세포
글리아 세포의 일종인 '성상교세포(astrocyte)'가 신경 전달 물질인 '글루탐산(glutamic acid: 20가지의 단백질 구성 아미노산 중의 하나)'을 받으면, 세포 속에서 칼슘 이온의 농도가 급격히 높아진다고 한다. 글리아 세포는 전기적이 아닌 화학적인 흥분을 일으키는 세포였던 것이다. 최근에는 여러 개의 성상교세포 사이에서 칼슘 사이에서 칼슘 이온의 물결이 전달된다는 점뿐만 아니라, 성상교세포의 흥분이 뉴런의 흥분까지도 유발한다는 사실이 알려졌다. 뇌 속에 퍼져있는 성상교세포의 네트워크가 뉴런의 네트워크와 서로 '정보(information)'를 교환하고 있었던 것이다.