'태반'의 과학
0. 목차
- '태반'은 어떻게 생겨났는가?
- 인간의 태반은 500g이나 된다.
- '탯줄' 안에 3개의 혈관이 나선 모양으로 꼬여 있다.
- '태반'은 물질을 주고받는 관문의 역할을 한다.
- '태반'과 '면역'
- '태반'을 만드는 유전자

1. '태반'은 어떻게 생겨났는가?
'태반(Placenta)'이란 폐·소화관·신장을 합친 것과 같은 기능을 하며, '기한 한정'으로 만들어져 출산과 함께 사라지는 장기이다. 태반은 기본적으로 모체와 태아를 연결해 산소를 보내고 이산화탄소를 모체로 보내며, 태아에게 영양분을 공급하고 노폐물을 모체로 보내는 기능을 담당한다. '태반'을 거대한 '내분비 기관'으로 파악하는 연구자도 있다. 임신을 유지하기 위해 필요한 여러 가지 호르몬, 예컨대 '인간 융모성 생식샘 자극 호르몬(hCG: Human Chorionic Gonadotropin)', '프로게스토론(Progesterone)', 젖샘을 자극하는 '인간 태반성 락토겐(hPL: Human Placental Lactogen)', '에스트로겐(Estrogen)' 등을 대량으로 분비하는 기능을 가지고 있기 때문이다.
1-1'태반을 가진 동물'과 '태반을 갖지 않은 동물'
'태반'은 임신했을 때 자궁안에 형성되는 기관이다. 그 조직은 스펀지처럼 되어 있으며, 산소와 영양을 풍부하게 함유한 어머니의 혈액으로 채워져 있다. 거기에서 자궁 내부를 향해 '제대(Umbilical Cord, 탯줄)'라는 혈관이 뻗어, 태아의 복부와 연결되어 있다. 배꼽은 출산할 때, 붙어 있던 탯줄이 떨어지면서 남은 흔적이다.
'태반을 가진 동물'은 거의 포유류에 국한되어 있다. 포유류는 '유성 생식으로 새끼를 낳고 젖으로 키우는 생물'의 총칭으로 약 4500종이 현존한다. 그리고 이들은 크게 세 그룹으로 나뉜다. '오리너구리', '가시두더지' 등의 '단공류(Monotremata)', 캥거루와 코알라 등의 '유대류(Marsupialia)', 인간과 생쥐 등의 '진수류(Eutheria)'이다. 약 1억 6600만 년 전에 세 그룹의 공통 조상으로부터 '유대류와 진수류의 공통 조상'이 갈라져 나왔다. 그리고 1억 4800만 년 전에 '유대류와 진수류의 공통 조상'에서 '진수류의 공통 조상'이 갈라져 현재에 이르렀다.

1-2. 태반의 진화
- 단공류(Monotremata): '단공류(Monotremata)'는 포유류 가운데 가장 원시적인 동물로, 알을 낳고 약 10일간 품어 부화한 새끼를 젖으로 키운다. 단 모체에는 젖꼭지가 없어, 젖은 복부 전체에서 스며 나온다. 알은 조류의 알과 거의 같은 구조이다. 발생 도중의 '배(Embryo)'는 '양막(Amnion, 태아를 둘러싸고 있는 얇은 막)'으로 감싸여 있으며, 복부에서 '난황낭(Saccus Vitellinus)'과 '요막(Allantois)'이라는 2개의 주머니가 나와 있다. '난황낭'에는 영양분이 채워져 있어, '배'는 여기서 영양을 받는다. '배'에서 생긴 노폐물은 '요막'에 고인다. 산소는 알껍데기를 통해 직접 받아들이며 배에서 생긴 이산화탄소도 껍데기를 통해 나간다.
- 유대류(Marsupialia): 다음에 출현한 '유대류(Marsupialia)'는 모체의 자궁안에서 자라지만, 진수류에 비하면 '난황낭 태반'이 얇고 발달이 덜되었다. 따라서 유대류는 아주 작고 미성숙 상태로 태어나며, 상당 기간 복부에 있는 주머니인 '육아낭(Marsupium)' 속에서 자란다. 예를 들어 캥거루는 새끼가 산도에서 육아낭까지 기어 이동하고, 육아낭 안에 있는 젖꼭지를 빨면서 그 상태에서 성장한다.
- 진수류(Eutheria): 마지막으로 등장한 '진수류(Eutheria)'는 '유태반류(Placentalia)'라고도 하며, 발달한 두꺼운 '융모막 태반'을 갖는다. 이러한 태반은 모체 안에서 시간을 들여 태아를 크게 성장시킬 수 있다. 태어난 새끼는 필요할 때 어머니의 젖을 먹으며 자란다. 한편, 진수류의 '융모막 태반'은 '요막'에서 유래한다. 인간의 경우 발생 7일째 무렵에 수정란의 일부가 '양막(자궁 내막)'의 세포와 강하게 접착해, 두껍게 발단한 태반이 만들어진다.
태아에게 영양을 공급해 키운다는 점에서, 태반은 '난황낭이 변해 모체 안에 달라붙을 수 있게 된 것'이라고 이해할 수 있다.
1-3. '태반'을 가진 동물은 공룡 멸종 후에 번영하였다.
'태반을 가진 생물'은 조류나 단공류보다 성숙한 상태로 새끼를 낳을 수 있게 되었지만, 그 이점이 발휘된 것은 공룡이 멸종한 약 6600만 년 전 이후의 일이다. 원시적인 '진수류'는 현존하는 생쥐보다 작고 야행성이었다고 한다. 공룡이 멸종한 이유는 거대한 소행성이 지구와 충돌하고, 그 이후의 기후 변동으로 기온이 낮아졌기 때문이라고 생각된다. 이러한 상황에서 공룡은 기후 변동에 적응할 수 없었다는 설이 유력하지만, 그 밖의 다른 종이 어떻게 살아남았늦지는 정확히 밝혀지지 않았다.
공룡들이 사라지자 '진수류'의 발달한 태반이 위력을 발휘하게 되었다. 알처럼 외적에게 먹힐 염려도 없고, 어미가 계속 알을 품을 필요도 없다. '생태적 지위(Ecological Niche)'를 넓힌 진수류는 다양해지면서, 마침내 대형이고 '주행성(낮에 활동하는 성질)'인 종도 나타났다. 진수류는 번영하면서 약 600만 년 전에는 침팬지와 인간의 공통 조상이 나타났으며, 약 20만 년 전에는 우리 현생 인류가 탄생하게 되었다.
2. 인간의 태반은 500g이나 된다.
2-1. 태반은 모양으로 보면 크게 4가지 형태로 구분된다.
이제부터는 인간의 태반에 대해 알아보자. '인간의'라고 제한한 이유는 태반의 구조와 구성 세포는 생물종에 따라 차이가 매우 크기 때문이다. 예컨대 진수류의 태반은 모양으로 따라 크게 4가지 형태로 구분된다. 자궁 일부에 둥근 원반처럼 만들어지고 침윤성이 가장 높은 '반상 태반(Discoid Placenta)', 양막의 중앙을 둥글게 감싸듯이 만들어지고 그다음으로 침윤성이 높은 '대상 태반(Zonary Placenta)', 자궁 안에 여기저기에 만들어지고 침윤성이 가장 낮은 '산재성 태반(Diffuse Placenta)', 양막에 산재하는 돌기 모양에 부착해 조그만 태반이 분포하는 '총모성 다태반'이다. 영장류와 설치류는 '반상 태반'을 가지고 있고, 개와 고양이는 '대상 태반'을 가지고 있으며, 말과 돼지는 '산재성 태반'을 가지고 있고, 소와 염소 등은 '총모성 태반'을 가지고 있다.
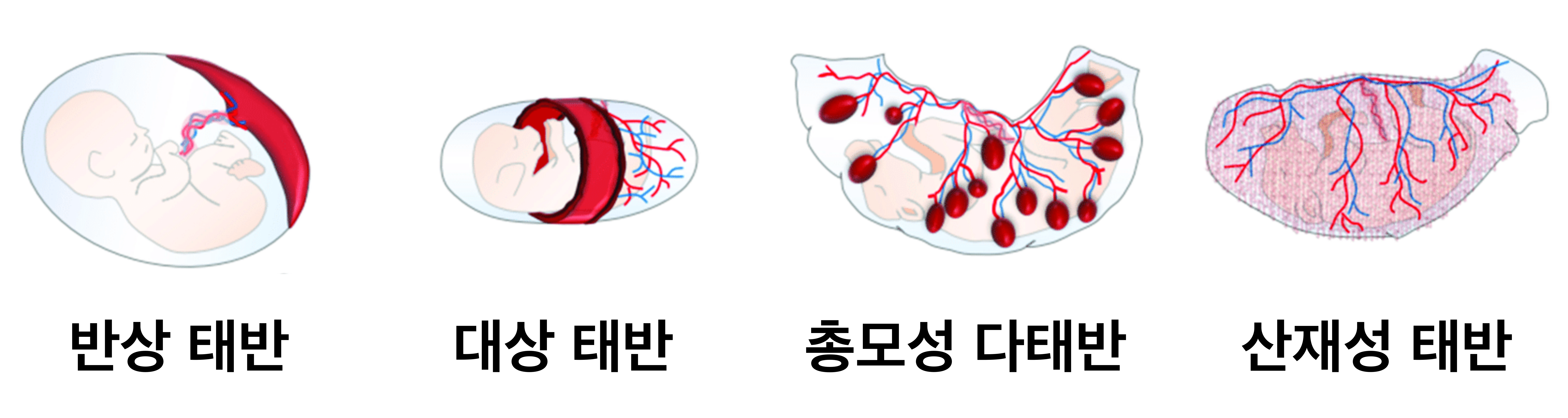
| 태반 | 태반의 모양 | 동물 |
| 반상 태반 | 자궁 일부에 둥근 원반처럼 만들어짐 | 인간, 생쥐, 토끼 등 |
| 대상 태반 | 양막의 중앙을 둥글게 감싸듯이 만들어짐 | 개, 고양이 등 |
| 산재성 태반 | 자궁 안 여기저기에 만들어짐 | 말, 돼지, 돌고래 등 |
| 총모성 다태반 | 양막에 조그만 태반이 분포함 | 소, 염소 등 |
2-2. 태반의 형성
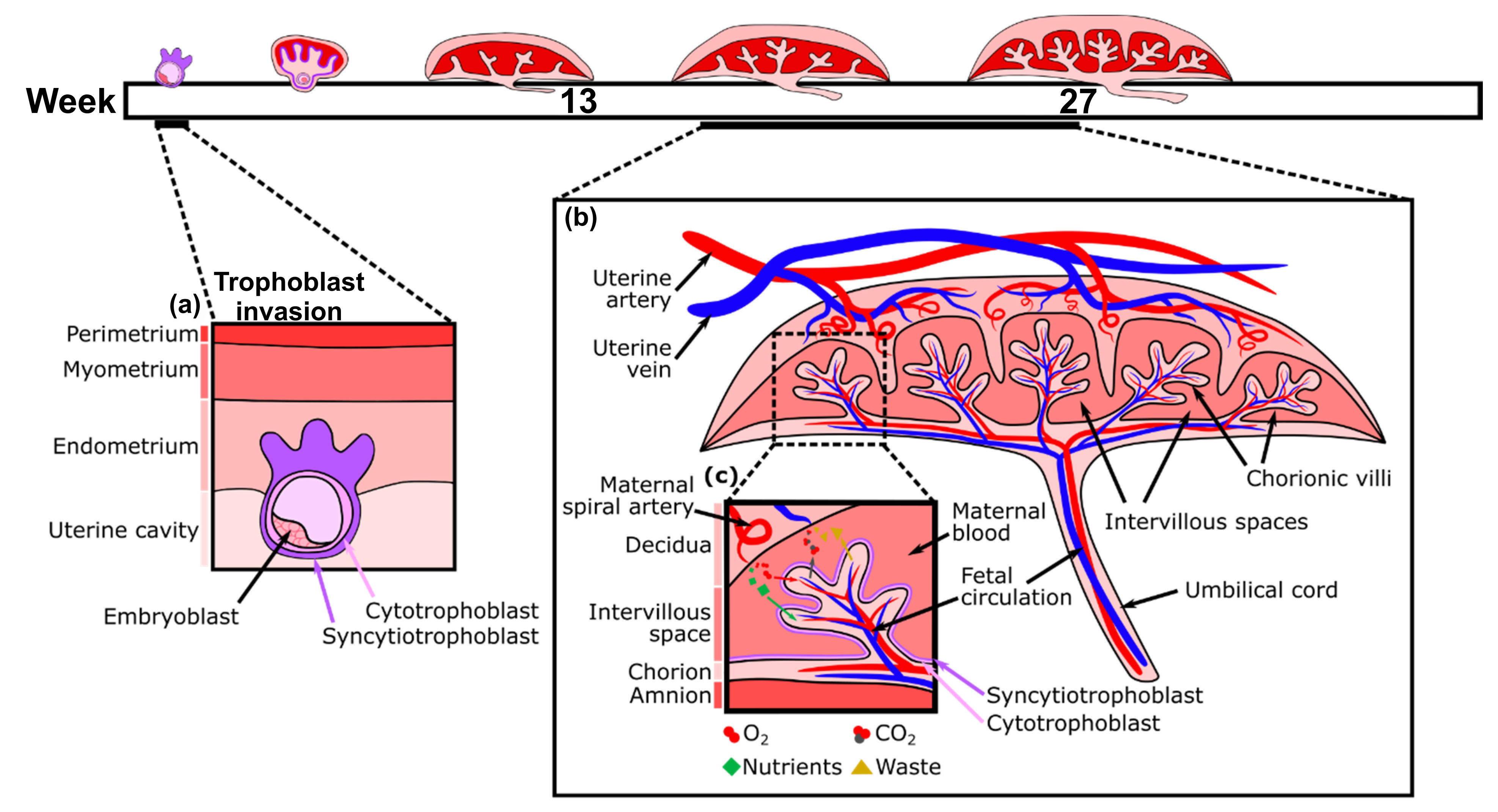
지름 0.1mm 정도인 인간의 수정란은 '양막(자궁 내막)'에 부착해, 약 36~40주에 걸쳐 몸길이 약 50cm, 무게 약 3kg의 태아로 자라난다. 그 성장을 지원하는 '반상 태반'은 임신 7주 무렵부터 만들어지기 시작해 16~20주까지 완성된다. 그리고 태반 구조가 아직 미숙한 임신 초기에는 어머니의 혈액 일부도 '배(Embryo)'로 옮겨지는 것으로 생각된다. 예전에는 자궁안에 생긴 태반은 '모체 쪽의 혈액으로 만들어진다'고 오해받기도 했지만, 태반의 대부분은 수정란의 일부, 즉 태아 쪽 세포로 만들어진다. 수정란에서 유래한 특수한 세포인 융모 세포가 자궁의 조직인 '기저 탈락막(Basal Decidua)'으로 들어가 만들어진 것이 태반이다.
'융모(Villus)'와 '기저 탈락막' 사이에 생긴 공간인 '융모간공(Intervillous Space)'은 모체의 혈액으로 채워져 있다. 융모 조직에는 태아 쪽의 모세 혈관이 감싸여 있어, 흘러들어온 모체의 혈액으로부터 산소와 영양을 받아들인다. 모세 혈관은 탯줄로 이어지고 다시 태아의 복부와 연결되어 있다. 태아의 모세 혈관은 얇은 '영양막(Trophoblast)'으로 덮여 있기 때문에, 융모 조직 안에서 모체의 혈액과 태아의 혈액은 직접 섞이지 않는다.
태반은 임신기를 지나면서 커져, 개인차는 크지만 출산할 때는 대략 지름이 약 20cm, 두께 몇 cm, 무게 약 500g의 원반 모양이 된다. '융모간공'에 모체의 혈액이 흐르기 시작하는 것은 임신 12주 정도 이후이며, 이 무렵부터 태아와 모체 사이의 물질 교환이 활발해진다. 임신 10주 무렵이 되면 모체에서 태반으로 이어진 나선 동맥 안의 세포가 태아 쪽의 세포로 바뀐다. 이러한 변화로 인해, 혈관은 지름이 약 10배 커져 모체로부터 대량의 혈액을 받아들일 수 있게 된다. '나선 동맥'은 임신 16주 무렵에 완전히 태 아쪽 세포로 바뀌며, 이를 계기로 태반이 완성되었다고 생각하는 것이 일반적이다. 태반은 출산 후 바로 떨어져 모체 밖으로 배출되어 짧은 기간의 역할을 마친다.
3. '탯줄' 안에 3개의 혈관이 나선 모양으로 꼬여 있다.
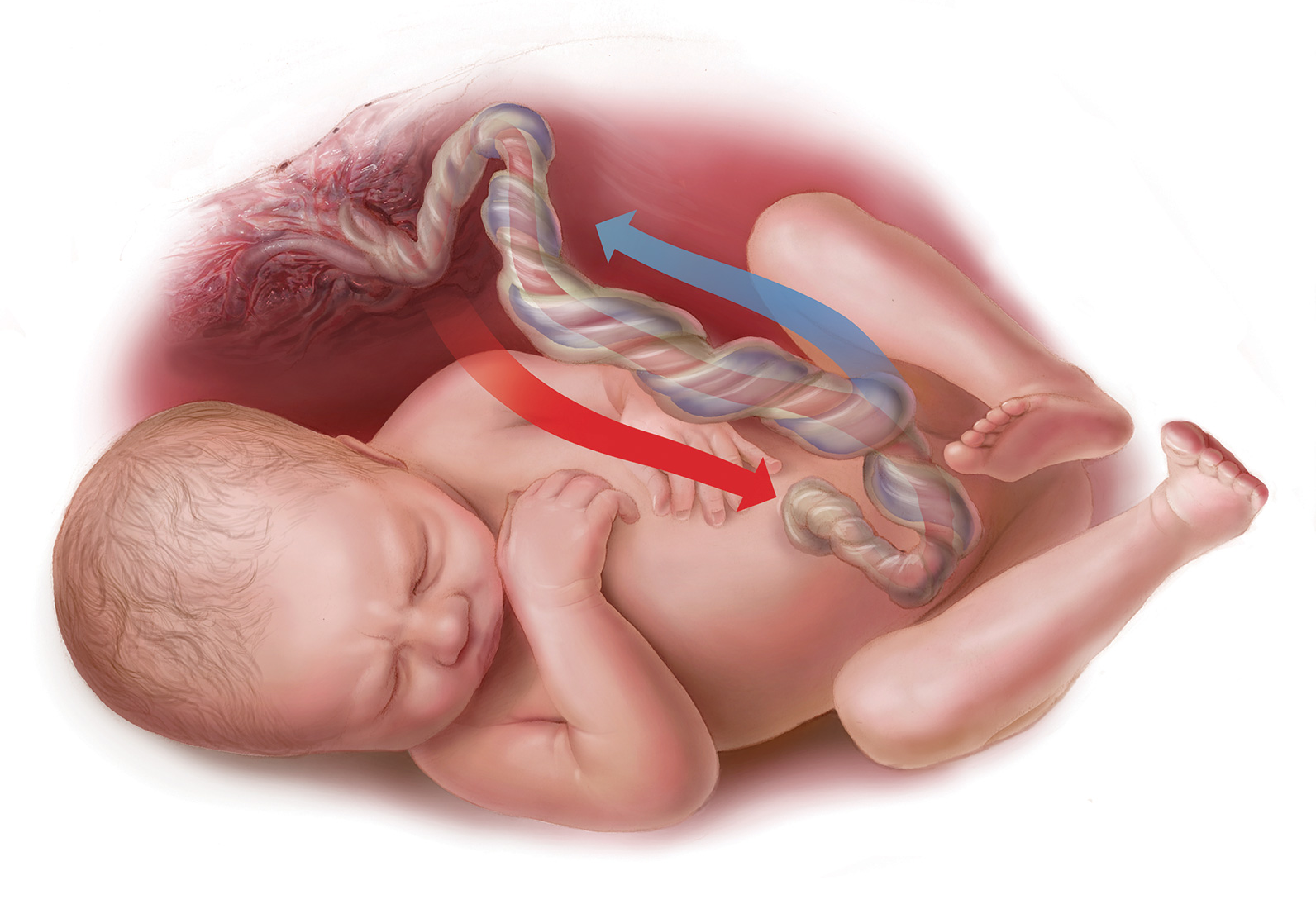
모체와 태아를 잇는 '제대(탯줄, Umbilical Cord)'의 구조도 매우 흥미롭다. 출생할 때의 탯줄은 길이가 약 50~60cm, 굵기가 약 2cm이다. 하얀색의 조직인 '와튼 젤리(Wharton's Jelly)'로 이루어진 이루어진 하나의 관 안에, 굵은 '제대 정맥'과 '약간 가는 2개의 ' 제대 동맥'이 들어 있다. 탯줄은 꼬인 나선 모양을 하고 있으며, 일반적으로 꼬이는 방향이 정해져 있다. 왜 꼬이는지는 밝혀지지 않았다. 나선 구조가 3개의 혈관을 간결하게 모아, 태아의 움직임에 따라 끌어당겨져도 부서지지 않도록 기능한다고 생각된다. 단 너무 단단히 꼬이면 혈류량이 줄어, 태아에게 발육 부전이나 사산 등의 '악영향'을 미친다. 약하지도 않고 너무 강하지도 않은 적당한 꼬임이 중요하다.
일반적으로 산소가 많이 포함된 혈액을 운반하는 혈관이 '동맥', 이산화탄소가 많은 혈관이 '정맥'으로 이해되는 경우가 많다. 하지만 태반에서는 반대이므로 주의가 필요하다. '제대 정맥'을 흐르는 것은 '산소와 영양을 많이 포함한, 태반에서 태아 쪽으로 흐르는 혈액'이며, '제대 동맥'을 흐르는 것은 '이산화탄소와 노폐물을 많이 포함한, 태아에서 태반으로 흐르는 혈액'이다.
'제대 정맥'에서 오는 혈액을 심장으로 순환시키는 '태아 순환계(Fetal Circulatory System)'는 태아기에만 존재하는 것으로, 하나의 혈관으로 이루어져 있다. '심장의 구멍'은 심장의 부담을 줄이고, 뇌에 우선적으로 혈액을 보내기 위해 있다고 생각된다. '심장의 구멍'은 출생 후에는 자연스럽게 막힌다. 태아의 몸속을 돈 혈액은 '제대 동맥'을 거쳐 태반으로 돌아온다. '제대 동맥'은 태아의 양쪽 발에 있는 혈관에서부터 갈라져 있다. '제대 정맥'은 하나이지만 '제대 동맥'이 둘인 이유는 태아의 이러한 사정에 의한 것이다.
4. '태반'은 물질을 주고받는 관문의 역할을 한다.
모체의 혈액에 포함된 물질은 태반의 '융모 조직'을 지날 때 취사선택된다. '관문'이라고 할 수 있는 이러한 기능을 담당하는 것은 성질이 다른 3종의 융모 세포가 융합한 '합포체 영양 세포막(Syncytiotrophoblast)'이다. 이 세포를 통과하기 쉬운 것은 '분자량이 작은 것', 혈장 단백질과 결합하지 않은 것', '지용성인 것', '이온이 되기 어려운 것'이지만 예외가 많다. 통과하기 어려운 물질이라도 태아의 성장에 필요한 것은 특수한 메커니즘이 작동해 통과하기 때문이다. 수송 방법은 크게 '수동 수송', '능동 수송', '수용체에 의한 수송' 3가지로 나뉜다.
- 수동 수송: 첫째는 물질이 농도 차이에 따라 높은 쪽에서 낮은 쪽으로 이동하는 '수동 수송'에 의한 것이다. '산소', '이산화탄소', '물', '전해질(물에 녹은 물질)' 등은 이 방법으로 수송된다. 예컨대 산소는 '산소 농도가 높은 모체의 혈액'에서 '산소 농도가 낮은 태아의 혈액'으로, '농도가 높은 태야의 혈액'에서 '농도가 낮은 모체의 혈액'으로 운반된다.
- 능동 수송: 둘째는 에너지를 사용해 세포막에 갖추어진 수송 도구 '트랜스포터(Transporter)'를 움직여 수송하는 '능동 수송'이다. 예컨대 '포도당(Glucose, 글루코오스)'은 수동 수송으로도 운반되지만, 태아에게 특히 중요한 물질이기 때문에 '합포체 영양 세포막(Syncytiotrophoblast)'의 '글루코오스 트랜스포터(Glucose Transporter)'에 의해 더 적극적으로 태아 쪽으로 공급된다. 단백질이 재료인 '아미노산'에도 각각의 트랜스포터가 있어 적극적으로 수송되도록 만들어져 있다.
- 수용체에 의한 수송: 셋째는 '세포막에 묻혀 있는 특수한 수용체'를 매개로 수송되는 것이다. 예컨대 병원체에 결합해 무력화하는 항체 '면역 글로불린G(IgG: Immunoglobulin G)'는 '특정 수용체(FcRn)'에 결합하면 통과할 수 있다. 외부의 병원체와 접한 적이 없는 신생아가 생후 3~6개월 사이에 감염증에 걸리지 않는 이유는 이 메커니즘에 의해 모체의 항체를 물려받기 때문임이 분명하다.
5. '태반'과 '면역'
5-1. 모자간 면역 관용
우리 몸의 면역 체계는 '자기(Oneself)'와 자기가 아닌 '비자기(Nonself)'로 구분한다. 그런데 난자와 정자가 만나서 만들어진 수정란은 모체 입장에서는 '자기'가 아니라 절반은 자기가 아닌 '반비자기(Half Nonself)'의 존재이다. 우리 몸에서는 병원체 같은 '비자기인 이물질'이 침입하면 그것을 물리치는 면역이 작용한다. 즉 '반비자기'인 태아와 태반도 면역 세포의 공격 대상이 되어야 하지만 그렇지 않다. 태반에는 태에서 유래한 물질은 이물질로 인식시키지 않은 특별한 메커니즘인 '모자간 면역관용(Immune Tolerance)'이 갖추어져 있다.
'모자간 면역 관용'을 이해하기 이해서는 '면역(Immune)'의 기본 원리를 알아야 한다. 과거에 감염된 적 없는 병원체가 몸속으로 침입하면, 처음에는 병원체의 종류와 관계없이 작용하는 '자연 면역'이 작동한다. 식세포라는 면역세포가 병원체를 닥치는 대로 집어삼켜 분해한다. 이러한 식세포의 일부는 병원체의 정보를 다른 면역 세포에 전달해, 더욱 강력하게 병원체를 물리치는 '획득 면역'이 발동하게 한다. '획득 면역'에서 병원체의 정보를 받은 'T세포'와 'B세포'가 협력해 작동하며 B세포가 만들어 낸 '항체'로 무력화하거나 T세포가 감염 세포를 공격해 병원체를 물리친다.
이러한 '면역 세포'는 모체의 혈액을 매개로 태반으로도 운반되는데, 태아 쪽의 세포에서 만들어진 '반비자기(Half Nonself)'인 융모 세포는 교묘한 메커니즘으로 면역 세포의 감시를 벗어난다. 우리 세포의 표면에는 자기임을 나타내는 표지인 'MHC 항원'이 붙어 있는데, 태반의 세포에는 이 표지가 없다. 즉, 융모 세포는 '면역계를 벗어나기 위한 물질(PD-1과 PD-L1 등)'을 만들어 낸다. T세포와 B세포는 '융모 세포'를 비자기로 인식할 수 없을 뿐 아니라, 활성화도 강하게 억제된다.
'모자간 면역 관용'은 태아가 성장하기 위한 매우 중요한 매커니즘이다. 정상적으로 기능하지 않으면 유산을 반복하거나 '임신 고혈압 신증'에 걸릴 위험이 높아진다.
5-2. '태반을 통과하는 병원체'와 '태반을 통과하기 어려운 병원체'
'어머니로부터 유래한 IgG가 태아에게 전달되는 메커니즘'과 '모자간 면역 관용'이 정상적으로 기능해도, 어머니가 임신 중에 감염증에 걸린 경우에는 태반과 태아 몸속에서 병원체가 증식해 강한 면역 반응이 일어나는 경우가 있다. 이러한 경우에는 일반형인 'NK세포(병원체에 감염된 세포를 공격하는 면역 세포)'가 증가해 병원체를 공격한다. 일반형 NK세포는 태반과 자궁에 염증을 일으키기 때문에, 많은 경우 태아는 살아갈 수 없어 유산된다.
단, 어머니가 감염된 병원체 모두가 태반을 통과하는 것은 아니다. 세균과 바이러스의 종류에 따라 '태반을 통과하기 쉬운 것'과 '태반을 통과하기 어려운 것'이 있다. 이유는 밝혀지지 않았지만 '사이토메갈로 바이러스(Cytomegalovirus)', '풍진 바이러스(Rubella Virus)', '에이즈 바이러스(Human Immunodeficiency Virus)', 'B형간염 바이러스(Hepatitis B Virus)' 등은 태반을 통과하기 쉽고, 통과하더라도 염증을 잘 일으키지 않는 것으로 알려져 있다. 따라서 유산하는 경우는 적지만, '사이토메갈로 바이러스'에 감염되면 발육 지연과 뇌신경계 이상, 그리고 '풍진 바이러스'에 감염되면 심장의 기형이 일어나는 경우가 있다.
6. '태반'을 만드는 유전자
이제부터는 '태반을 만드는 유전자'의 기묘한 작동 방식과 그 유전자가 어떻게 생물의 '유전 정보(Genome)'와 관련되는가라는 점에 주목해 보자.
6-1. 태반을 만드는 유전자 PEG10
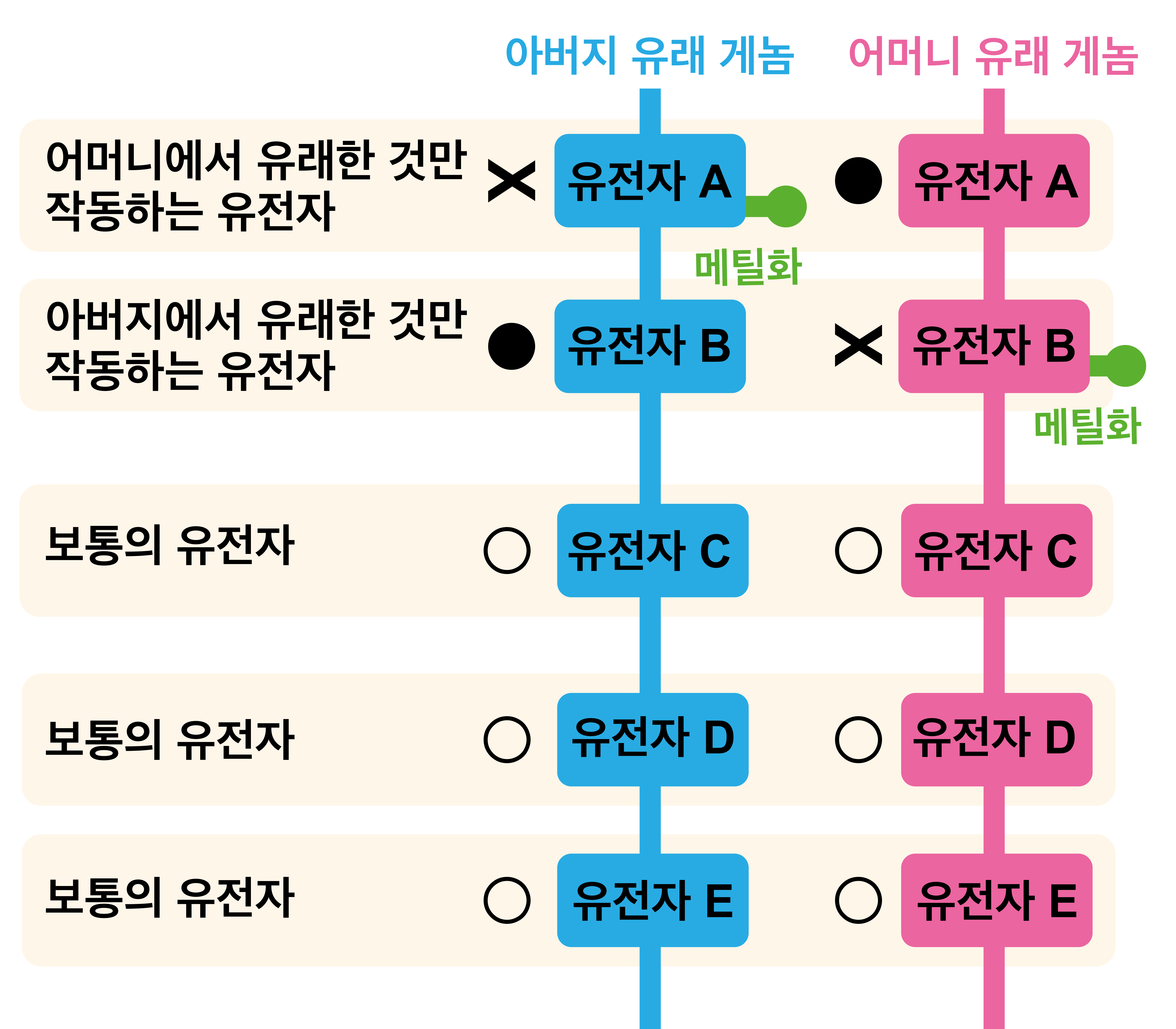
인간의 세포에는 23종의 염색체가 2세트, 합계 46개가 있다. 부모로부터 1세트씩 물려받은 것이다. 그런데 배의 발생 과정에서 '어머니 유래'와 '아버지 유래'의 어느 한쪽의 유전자가 중요한 기능을 하는 것으로 알려져 있다. 보통의 유전자는 아버지에게서 유래한 것과 어머니에서 유래한 것이 동등하게 있지만 예외가 있다. 사용되지 않은 쪽에는 '유전자를 기능하도록 하기 위한 DNA 배열(제어 영역)'에 '메틸화'라는 화학적인 작용이 가해져 기능하지 않게 된다. 이처럼 한쪽의 유전자가 '메틸화'되어 기능하지 않아, 한쪽의 유전자만 작동하는 유전자를 '임프린트 유전자(Imprint Gene)'라고 한다.
예컨대 아래 그림에서 유전자 A와 유전자 B는 '임프린트 유전자'이다. 도쿄대의 '이시노 사시토시' 교수가 우연히 발견한 '태반을 만드는 유전자'도 '임프린트 유전자' 가운데 하나이다. '임프린트 유전자'는 현재까지 수십 개 이상 발견되었으며, 발생 초기에 '아버지 발현 임프린트 유전자(아버지에서 유래한 것)' 또는 '어머니 발현 임프린트 유전자(어머니에서 유래한 것)'이 작동하도록 제어된다.
1980년대 초에 생쥐 수정란의 6번 염색체의 특정 부위를 '아버지에서 유래한 것'이 아니라 '어머니에서 유래한 것'으로 바꾸자, '배(Embryo)'가 자라지 못하고 죽은 경우가 보고되었다. 연구진들은 이 영역에 태반 형성에 '중요한 아버지 임프린트 유전자'가 있다고 생각해 탐색을 계속하다가, 2003년에 '아버지 발현 임프린트 유전자'의 하나인 PEG10 유전자를 찾아냈다. 그리고 분석을 계속해 정상적이 생쥐의 태반에서는 PEG10 유전자가 매우 많이 작동한다. '아버지에서 유래한 PEG10 유전자가 작동하지 않도록 유전자를 조작한 수정란은 태반이 제대로 만들어지지 않아, 태아는 발생 9.5일 이후에 죽고 말았다'는 사실이 밝혀졌다.
6-2. PEG10 유전자는 레트로바이러스 감염에서 유래했다?
'이시노 사시토시' 교수 등은 기존의 게놈 배열을 사용해, 어떤 생물종이 PEG10 유전자를 가지고 있는지 조사했다. 그 결과, 태반을 가진 모든 생물종에는 공통적으로 PEG10 유전자가 있으며 '단공류', '조류', '어류' 등의 난생 생물에는 없음을 확인할 수 있었다. 인간의 경우 7번 염색체에 PEG10 유전자가 있다. PEG10 유전자의 발견과 같은 시기에, 태반이 발달한 '진수류'에게만 보이는 또 다른 '아버지 발현 임프린트 유전자' PEG11 유전자를 찾아냈다. PEG10 유전자와 PEG11 유전자에 의해 만들어진 단백질인 아미노산 배열을 상세히 검토한 결과, 이들이 '레트로트랜스포존(Retrotransposon)'이라는 배열에서 유래한다는 사실도 밝혀냈다.
'레트로트랜스포존(Retrotransposon)'이란 자신을 복제해 게놈 여기저기에 삽입하는 특수한 DNA 배열의 하나로, 원래는 '레트로바이러스의 유전 정보의 일부였던 것'으로 생각된다. '레트로바이러스(Retrovirus)'는 유전자로서 RNA를 가지며, '역전사 효소(Reverse Transcriptase)'를 사용해 RNA에서 DNA를 만드는 바이러스의 총칭이다. '레트로 바이러스'는 '역전사 효소'를 사용해 RNA로부터 DNA를 만들고, 그 일부를 감염되는 세포의 DNA에 삽입하는 특징이 있다. '레트로바이러스'에는 '인체 면역 결핍 바이러스(HIV)'와 '인간 T세포 백혈병 바이러스(Human T-Cell Lymphotrophic Virus)' 등이 있다.
6-3. '난황낭'과 '요막'은 융모 세포를 매개로 모체의 내부에 달라붙게 되고 그것이 '태반이 되었다?
이에 '이시노 사시토시' 교수는 PEG10 유전자의 바탕이 된 '레트로트랜스포존'도 레트로바이러스 감염에서 유래한 것이라는 가설을 세웠다. 그 가설은 다음과 같다. 1억 6600만 년 전보다 이전에 '난생 동물(알에서 새끼가 나오는 동물)'이 어떤 레트로바이러스에 감염되었다. 그때 바이러스의 유전 정보의 일보가 세포의 DNA 안으로 삽입되었다. 삽입된 DNA 배열은 '생식 세포(난자와 정자)'를 매개로 당음 세대에 전해졌다. 세대를 거치는 사이에 DNA 배열 안에서 돌연변이가 축적되고 마침내 PEG10 유전자로 변했다.
그리고 약 1억 6600만 년 전, 우연히도 PEG10유전자는 수정란의 '난황낭(Saccus Vitellinus)'과 '요막(Allantois)'에 접착성이 강한 특수한 세포인 '융모 세포'를 만드는 기능을 갖게 되었다. '난황낭'과 '요막'은 융모 세포를 매개로 모체의 내부에 달라붙게 되고 그것이 태반이 되었다. 그 후 같은 방식으로 PEG11 유전자를 획득한 것이 나타나 '더욱 발달한 요막 유래 태반' 기능을 장기간 유지할 수 있는 '진수류(Eutheria)'가 되었다.
부모 어느 한쪽의 유전자만을 사용하는 '게놈 임프린팅(Genome Imprinting)' 현상은 태반을 가진 '유대류(Marsupialia)'와 '진수류(Eutheria)'에서만 볼 수 있다. '게놈 임프린팅'은 바이러스 감염 등에 의해 삽입된 외래 배열의 기능을 억제하는 기능과 관계있을지도 모른다.
6-4. 바이러스에 감염되어 게놈은 진화를 계속한다.
레트로바이러스의 감염이 유래라고 생각되는 DNA의 배열은 생물의 '게놈(Genome)' 곳곳에 남아 있다. 바이러스는 지금도 게놈 속으로 들어오려 하고 있다. 예컨대 2000년 이후에 코알라에서 감염이 확대되고 있는 '코알라 레트로바이러스(KoRV: Koala Retrovirus)의 유전자가 일부 이미 코알라의 게놈에 들어와 있음이 확인되었다. 게다가 코알라의 게놈에 삽입된 DNA 배열은 다음 세대에도 계승된 것으로 밝혀졌다. 게놈에 자리한 많은 DNA 배열 가운데, 다양한 변이를 거쳐 우연히도 유익한 단백질을 만들어 낸 것이 '그 생물의 유전자'로 보존될 것이다. 우리의 게놈은 바이러스에게 커다란 영향을 받아 함께 진화해 온 것이다.